Rho GTPase による ROS の制御
細胞内の ROS/RNS 生成は、主に、NADPH オキシダーゼ(NOX)、
一酸化窒素合成酵素(NOS)、
スーパーオキシドジスムターゼ(SOD)などの、
主要な酸化還元酵素に由来します5,9,10(図1)。
Rho GTPase(特に Rac1と Rac2)は、
活性化されたNOX 複合体の構成要素であり、
続いて起こる酸素分子(O2)からの ROS(スーパーオキシドアニオン、O2.-)生成に必要となります11。
細胞外シグナルによって活性化されると、
NOX 複合体は細胞膜で会合し、Rac(GTP結合型、おそらくGDIが結合)の関与によりROS が生成されます12,13(図1)。
Rac が、アダプターとして作用するのか、
O2.- 生成の電子伝達を仲介するのかは、未だ明らかになっていません12。
また、Rac(特に Rac1)は、
一酸化窒素(NO)の生成を制御する酵素である NOS と、
直接相互作用することが示されています13。
NOS は、ヌクレオチドフリー型の GTPase に優先的に結合する
ことが報告されており、Rac が仲介する NO 産生に、
グアニンヌクレオチド交換因子(GEF)、
及び GEF を介した細胞内区画化が関与していることが示唆されます13。
さらに、Rac は、
O2.- の過酸化水素(H2O2)への変換を触媒する酵素である SOD と、酸化還元によって直接相互作用することが示されています14。
還元条件下では、細胞内と同様に、
Rac-GTP が SOD に結合することで Rac の活性が維持され、
NOX による O2.- の産生が促進されます。
酸化条件下では、局所的な O2.- / H2O2 生成と同様に、
まだ同定されていない GTPase 活性化タンパク質(GAP)
によって、SOD が Rac から解離し、不活性型である GDP 結合型に変換されると考えられています14。
これらの報告から、Rho GTPase は、
多数の活性酸素中間体(RIs)を生成し、
多様なタンパク質修飾をもたらす可能性があることが明確に示されます。
生成される ROS/RNS、及び誘発される細胞応答の種類は、
与えられた刺激、細胞内の局所的な酸化還元電位、細胞の位置などによると考えられます。
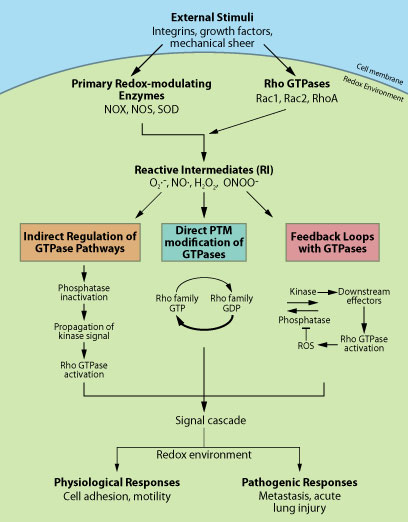
図1 シグナル伝達における
Rho GTPase - ROS 間のクロストーク
細胞内において想定される Rho GTPase - ROS/RNS 間の
クロストーク概略図。
間接的(オレンジ色)、直接的(青色)、
フィードバック(ピンク)に分類し、その下に、
最も一般的なメカニズムを示した。
各経路の詳細については、参考文献および本文中の例を参照。
ROS による直接的/間接的な Rho GTPase の制御
近年、 酸化還元を介した翻訳後修飾(PTMs)によって、
ROS/RNS が直接 Rho GTPase を制御する可能性がある、
という興味深い報告がなされています。
0 件のコメント:
コメントを投稿